Managing the equine estrous cycle is a common procedure performed by many broodmare managers and veterinarians. Copious research has elucidated many aspects of the equine estrous cycle and allowed veterinarians the tools to manage a mare’s cycle to provide the optimal breeding times, effectively use artificial insemination, induce ovulation, synchronize mares, induce superovulation, advance the onset of the breeding season, terminate pregnancy, as well as a manage a host of other reproductive conditions and diseases. In this article Dr. David Scofield reviews the use of progesterone and estradiol (P&E) therapy in the mare.
Before embarking on a discussion regarding cycle manipulation, an understanding of the interplay of the hypothalamic-pituitary-gonadal axis (HPG), various hormone release patterns, and overall follicular wave dynamics are a necessary pre-cursor. Mares are seasonally polyestrous animals that experience a period of winter anestrous, followed by a spring transition to cyclicity, regular estrus cycles during the long days of spring and summer, and a fall transition again to winter anestrous. During the physiological breeding season, only pregnancy, or in some cases, uterine or ovarian pathology can cause interruption of a mare’s 21-day inter-ovulatory window.
For the sake of this topic, we can assume the mare is regularly cycling and entered the breeding season. Entire textbooks are written on the physiology and endocrinology of the mare, and the following is a brief introduction to the cyclic mare. For previous discussions regarding advancing the start of the breeding season or managing the transitional mare, please see the blog post, Hastening the Onset of the Breeding Season.
The Estrus Cycle
A generic estrus cycle is divided into periods termed proestrus, estrus, metestrus, and diestrus. In the mare, however, we have generally condensed these into periods defined by either behavioral terms of estrus and diestrus or physiological terms of follicular and luteal phases. Two major steroid hormones are used to describe these two behavioral periods. The first is estrogen from the cells of the dominant follicle. Theca and granulosa cells respond to LH and FSH respectively to synthesize increasing amounts of estradiol that peaks near the time of ovulation. During the follicular phase/estrus, the dominant ovarian hormone is estradiol. During the luteal phase or diestrus, the steroid hormone produced by the corpus luteum is progesterone that prevents a mare from showing signs of estrus when in the presence of a stallion. These two follicular and luteal phases are intertwined however, because even though the luteal phase is dominated by the corpus luteum and progesterone production, there are still follicular wave dynamics and the recruitment and selection of the subsequent dominant follicle.
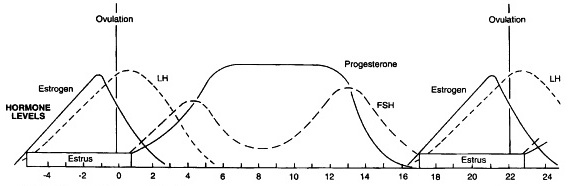
The pulsatile secretion of gonadotropin-releasing hormone (GnRH) from hypothalamic neurons in the median eminence to portal blood of the pituitary gland drives follicle stimulating hormone (FSH) and luteinizing hormone (LH) from the anterior pituitary gland.
LH and FSH control the selection, growth, maturation, and eventual ovulation of the dominant follicle.
Having one releasing hormone, GnRH, responsible for the secretion of two separate protein hormones from the pituitary having separate functions poses a problem for the maturing follicle. However, nature has developed a nice interplay between GnRH pulse patterns as well as ovarian feedback loops to affect the differential production and release of FSH and LH and help manage follicular dynamics.
In general, just after ovulation, there is a small rise in FSH that signals the recruitment of primordial follicles to begin a period of growth and maturation. This growth is FSH dominated during the luteal phase by slow hypothalamic GnRH pulses that favor FSH production in the pituitary. FSH drives the early stages of follicular maturation and peaks around days 10-12 of the cycle before declining. The decline in FSH is due in part to changes in the GnRH pulse pattern, but mostly due to negative feedback from rising inhibin and estradiol-17β (E2) production from the maturing follicles.
The maturing follicular wave will deviate at ~22.5 mm and a dominant follicle will grow at a rate of 3-5 mm a day until reaching a plateau prior to ovulation.
This dominant follicle secretes increasing amounts the protein hormone inhibin and steroid hormone estradiol-17β until just prior to ovulation. As the dominant follicle increases production of these two hormones, the remaining follicles in the wave become destined for atresia and regression.
Following deviation, the dominant follicle has increased sensitivity to the lower circulating levels of FSH. Final growth and maturation, however, is dependent upon increasing LH pulses and the high levels of LH observed during the LH surge. Faster hypothalamic pulses of GnRH favor LH subunit production and release from the pituitary, and the high levels of inhibin and E2 favor LH over FSH production. The specifics regarding the LH surge are beyond the scope of this blog, but the interplay between follicular wave dynamics and circulating hormones is crucial to understanding the following discussion on therapies.
P & E Therapy
Knowing the hormones involved and the pattern of negative feedback, we can attempt to exert some control over the follicular dynamics and cycle of the mare. To begin, we can look at the use of progesterone and estradiol (P&E) as therapy in the mare.
P&E has been used since the early 1980’s to “program mares,” synchronize mares, manage timed breeding, and to “restart” the cycle in mares with aberrant and irregular cycles.
By far the two most common uses of P&E is to program mares for a specific time when a stallion will be available and to synchronize mares for either embryo transfer or to maximize semen usage.
Looking at these hormones individually will demonstrate why P&E is effective. Although massively simplified, progesterone has a negative effect on LH and estradiol has a negative effect on FSH. One way to remember this fact, is that pregnant mares have high levels of circulating progesterone from ovarian sources, especially prior to day 100 of pregnancy. Their ovaries however, continue to have follicular waves during this period of time. High progesterone concentrations slow GnRH pulse frequency and the mimic mid-luteal GnRH slow-frequency pulses that favor FSH secretion by gonadotropes in lieu of LH. Although progesterone alone cannot control follicular waves, it does control estrus behavior and will cause preferential release of FSH over LH from the pituitary.
Estrogens, however, have a more complicated pattern of influence on the hypothalamus and pituitary gonadotropes. In other species, estrogens can have both a stimulatory or inhibitory effect of pituitary hormones depending upon the time and duration of administration. It is known in mares, that E2 administration will decrease FSH secretion and will control follicular wave dynamics. In mares, high circulating E2 will also increase the quantity of LH stored in secretory granules in the pituitary, helping to increase the release of LH in response to GnRH at the time of the LH surge.
Administration of 150 mg progesterone and 10 mg estradiol-17β for ten days will effectively control follicular waves.
This therapy prevents new follicular waves from developing past the 18-22 mm size. It does not, however, prevent existing pre-ovulatory follicles from ovulating and forming a CL. Since ovulation can occur during P&E therapy, a dose of prostaglandin or prostaglandin analog is added to the therapy on the final day of treatment. This 10-day protocol leaves ovaries with a cohort of 18-22 mm follicles that will begin maturation, deviation, and ovulation following withdrawal of P&E administration.
This protocol provides very consistent timing of ovulation. Most mares will ovulate 10-12 days following the last dose of P&E.
Early studies using this protocol had 13 of 16 mares ovulate on days 10-12 following the last dose of P&E. No ovulation induction medications were used in this study that could have provided a tighter degree of synchrony. This easy management tool allows stud farm and broodmare managers to manage the book of a busy stallion early in the breeding season.
Using hCG or deslorelin to control ovulation provides even tighter control of the mare’s ovulatory period following P&E.
Besides programming mares for stallion availability, P&E is the most effective way of synchronizing mare’s cycles for use as recipients in an embryo transfer program. Shortening a mare’s diestrus period by using prostaglandins is a common method utilized to line-up stages of the cycle in mares. This method, however, rarely produces the type of synchrony needed for use as a recipient. By starting mares on P&E therapy, the needed degree of synchrony can be accomplished.
Lastly, some practitioners use P&E to “reset” the cycle of a mare that has irregular or aberrant ovulations. Mares that routinely have diestrus ovulations that alter cycle length or failure to develop normal wave patterns of folliculogenesis can benefit from bringing the status of her ovaries back to a single size of follicle and letting this cohort mature from a common starting point.
In all cases, the use of P&E should be reserved for mares regularly cycling during the breeding season or for mares late-in-transition. P&E is good protocol for mare owners wishing to use their own recipients or stallion owners wanting to have an early season breeding plan for a heavily booked stallion. Formulations containing both progesterone and estradiol are available from compounding pharmacies. Check with your veterinarian regarding the specific use of this protocol for your horses and catered management for your reproductive needs.

